総合
要約
強迫症(ピッタリ系・確認系・洗浄系)は、CSTC回路の過剰に起因とする仮説が有力である。まず前頭前野で行動計画が作成されるが、それは不快感やエラー感などの情動面とそれに対処する行動面がミックスされたものであり、線条体と視床で情動面の興奮度が加算(ゲイン)されて戻ってくる。そのため、行動計画は2回目のループに入り、さらには、適切な抑制作用が無ければ無限ループに陥る。その過程で、行動面が習慣回路へと移行し、強迫観念(OFC–尾状核) × 強迫行為(背側被殻)の二重構造(回路)が生じる。この二重構造が強迫症の本質であり、二重の駆動力が個人の心身を蝕んでしまう。
各強迫症の発症原理は共通であるが、症状(ピッタリ系・確認系・洗浄系)に違いが出るのは、基本原理にプラスする作用点が違っているからである。本稿の主題であるピッタリ系では、PVニューロン(介在ニューロン)の障害により「補償的に幼児期の行動様式が優位になる」ことが特徴的である
第1章 強迫症発症の共通神経基盤
第1節 心理モードと最重要脳神経部位
(心理モードとは)
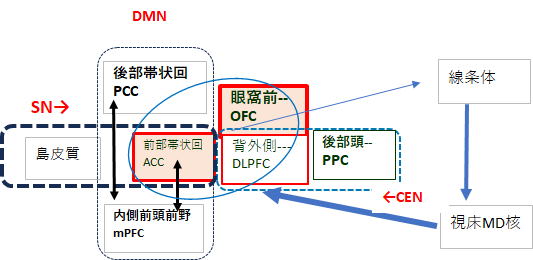
・デフォルトモード・ネットワーク(DMN) 何も外部の課題に取り組んでいない安静時や内省的な状態で活 動が高まる脳内ネットワークで、主な構成領域は内側前頭前野(mPFC)や後部帯状皮質(PCC)、楔前部
(Precuneus)などです。
・サリエンス・ネットワーク (SN) 前部島皮質(AI)や前帯状皮質(ACC)を中心とするネットワークで、外界から の刺激や体内からの信号の中から生存や目標達成に重要なものを選別し、注意を振り向ける役割を担います
・セントラル・エグゼクティブ・ネットワーク (CEN) 「集中・実行・意思決定」を担う脳の司令塔
ネットワークで、OFC(眼窩前頭皮質)や
DLPFC(背外側前頭前野)が中核領域です。
(CSTC回路の始点となる神経部位)
・OFC(眼窩前頭皮質) 報酬・罰・感情価値を評価し、「どれが良い選択か」を判断する。 情動・動機
づけ衝動の調整にも深く関わる。
・DLPFC(背外側前頭前野) ワーキングメモリ、計画、問題解決、抑制などの実行機能を担う。
論理的思考や目標達成のための行動制御を行う。
・ACC(前部帯状回) 情動・意思決定・注意・痛みなど多様な機能を担う重要領域
第2節 強迫症の前段階
強迫症と個人の「考え方の傾向」には密接な関係があり,責任の過大評価、不確実性への耐性の低さ、思考と現実の混同(考えただけで現実化する感覚)などが指摘されています。そのような考え方をベースに、ストレス・失敗体験・責任の重い環境などが引き金 となって強迫症が発症すると考えられています。
これらを脳神経レベルの用語で説明すると次のようになります。心理モード(DMN)が賦活して、後部帯状回と内側前頭前野の連絡が密になり内省的傾向が強まります。同時に心理モード(SN)も賦活して、前部帯状回と内側前頭前野の連絡も密になります。この状態は心理モードDMNとSNが前部帯状回を仲立ちとして、二つのギアの歯車がかみ合って回転し始めることを意味します。
一方で、OFC(眼窩前頭皮質) と心理モード(CEN)の中核DLPFC(背外側前頭前野)、ACC(前部帯状回)の三者は機能的結合が強まります。これは視床(MD)核の働きによるものです。この段階では個人の考え方の傾向(上記)は敏感さを増していき、何らかのきっかけにより、不安感に具体的な対象がセットされてしまいます。これは一つのギアに例えることができ、上記の心理モードのギアと歯車ががっちりと嚙み合ってあってしまい、結果、CSTC回路の果てしなき回転が始まってしまうのです。
第3節 線条体の機能
(PVニューロン)
さてこれからCTC回路のT(線条体)の解説にはいりますが、これはかなり複雑なので、順を追って説明します。ちなみに、線条体とは、大脳基底核を構成する中心的な領域で、尾状核と被殻から成り、運動・学習・意思決定・報酬処理に深く関わる脳部位のことです。
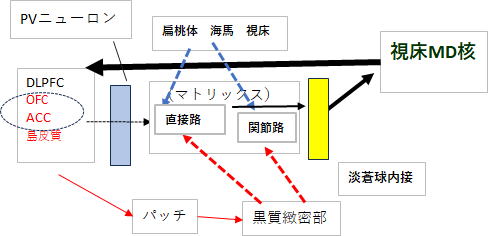
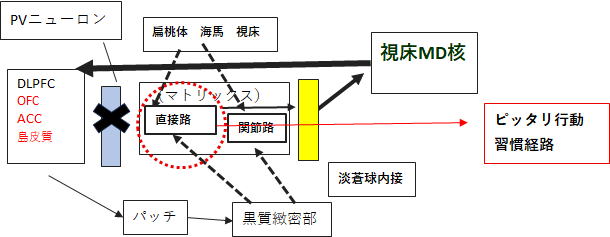
PVニューロンの左側に位置するのは、皮質(前頭前野)から送られてきた行動計画の構成要素です。各要素はそれぞれ独自かつ並列的に進んできます。中心はOFC(眼窩前頭皮質) ・DLPFC(背外側前頭前野) ・ACC(前部帯状回)ですが、他に島皮質も加わっています。OFC・ACC・島皮質は赤文字となっていますが、これらは、エラー感や不全感などの情動面を担当する構成部位で、DLPFCは具体的な行動計画の内容を含んでいます。
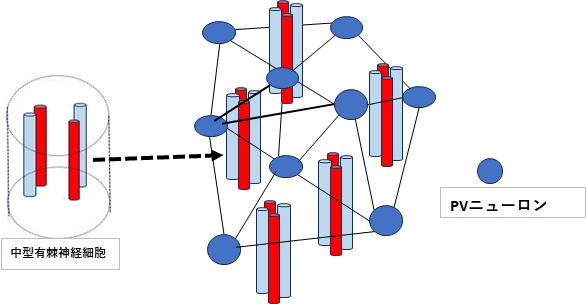
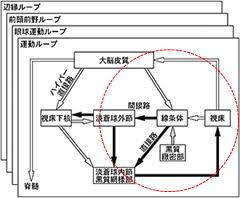
尾状核はパッチとマトリックスに分かれていますが、マトリックスの中にパッチが点在しています。瀬戸内海(マトリックス)、島々(パッチ)のイメージです。行動計画の赤文字部分は情動面を担当する神経部位で、これらはパッチに投射し、続いてパッチ→黒質緻密部→マトリックス→ドパミン投射という流れになります。マトリックス内にはgaba投射性の中型有棘神経細胞が存在し、ドパミンD1受容体を持つ直接路(MSN)と、D2受容体をもつ間接路(MSN)に分かれています。
(直接路と関節路)
この直接路(D1-MSN)と間接路(D2-MSN)は理解するのは多少ややこしいです。直接路は運動促進・行動選択の「GO」シグナルを強め、間接路は逆に抑制性の「NO-GO」シグナルを強めます。黒質緻密部からドパミン投射を受けると、直接路(D1-MSN)は発火率が上がり、運動促進・行動選択の「GO」シグナルを強めます。
それでは、間接路(D2-MSN)はどうなるか。この場合でも 運動促進・行動選択の「GO」シグナルを強めるのです。ここが理解しずらところですが、D2受容体は抑制性(マイナス)の効果をもたらすので、「NO-GO」のNOを抑制することになり、結局GOシグナルを強めます。そのため間接路が本来の意味を活かすのは、黒質緻密部からドパミン投射が極端にすくないケースで、文字どおり「NO-GO」になり行動は抑制されます。
線条体は、さらに、扁桃体・海馬・視床からもグルタミン酸投射を受けます。これにより直接路(D1-MSN)と間接路(D2-MSN)の膜電位は上がっていきます。
(PVニューロン)
線条体にはGABA投射性のPVニューロンが存在し、これらはMSNの発火タイミングを強く制御します。発火タイミングを制御するとは、PVニューロンが抑制性の神経伝達物質gabaを線条体MSNに広範囲に投射することによって、瞬間的にMSNの膜電位を下げることです。この直後の間隙をついて皮質からの行動計画が同期してMSNに投射することにより、MSNと皮質からの行動計画のタイミング(リズム)が一致して、行動計画が線条体に正しく投射されます。つまり、PVニューロンの働きの第一は「行動計画が正しく線条体へ通るための時間的ゲートを形成する」ことであり、結果として、両者の位相が疑似的に一致し、ノイズ(競合)を抑制することができます。
ところで、上記の記述はPVニューロンの機能を強調するために軽く流したところがあります。上記赤文字、皮質からの行動計画が同期してMSNに投射とありますが、この箇所(意味)は極めて重要ポイントです。ポイントは行動計画が同期してという部分です。 図2を見ていただければ分かりますが、PVニューロンの左側には皮質から並列的に投射された神経部位が並んでいます。 すなわち、DLPFC・
DFC・
ACC・島皮質です。 これら神経部位からの投射はそれぞれ周波数が違っています。この4者の周波数が、20hz・40hz・60hz・80hzであれば、この4者が同時に位相同期(リズムを合わせる)ことは簡単ですが、実際にはこのような数字配列にはならないので、4者が同時にリズムを合わせることは事実上不可能です。そのため最初は4者のうち2者が位相同期し、続いて3者、4者とリズムを合わせて、結果的に4者がリズムを合わせ同期した情報として線条体へと投射されます。
さて、行動計画は、等しく直接路と間接路へ投射されますが、どの神経細胞群のどの位置に投射されるか(空間的パターン)で情報は表現されます。また投射先のD1受容体とD2受容体の密度により、直接路と間接路の発火率が違ってきます。
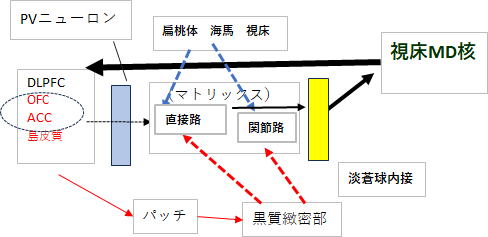
上記のシステムによって線条体を通過した行動計画は、淡蒼球内接で一つの内容に集約され、視床へ投射されて、他はノイズとして焼却されます。
選択基準は「直接路と間接路」そして「行動計画間」の相対評価です。CSTC回路は同時に複数の行動計画が並列的に走っています。これは上記の行動計画をAとすると他にもB・C・・の行動計画が存在していることを意味します。仮に行動計画が3本あるとすると、評価対象は、それぞれ「直接路・間接路」の2パターンを掛け合わせて、3×2=6 この6本の中から最も線条体MSN(中型有棘神経細胞)の興奮度(発火率)が高い内容が選択されます。そして生き残った計画は、淡蒼球内接→視床の抑制回路に抑制をかけ通行可能な回路の中を視床へと通過していきます。
しかし、この生き残り視床へと投射される行動計画は、最初の段階の行動計画の完全コピーではありません。黒質緻密部からドパミン投射を、また偏桃体・海馬・視床からグルタミン酸投射をうけて、選ばれて協調そして洗練されたバージョンになっています。
図2ではそれを細線→やや太線として表現しています。
第4節 視床の機能
視床には感覚中継核と連合核の両方が存在します。感覚中継核は視床に入ってくる感覚情報(嗅覚以外)を大脳皮質へ中継する核群。そしてもう一つが連合核で、前頭前野と連絡し、意欲・意思決定・社会的行動に関与 しています。CSTC回路に存在するのは連合核のうちの(MD核)です。
前頭前野は新脳、視床は旧脳に属していて、視床がより上位(管理者的立場)の前頭前野の機能を調整することに違和感がありますが、新脳と旧脳は互いに管理・調整する関係にあります。「互いの暴走を防ぐ仕組み」なのでしょうか?
視床
MD 核は「OFC・ACC・DLPFC の情報を選別し、同期させ、必要なネットワークを強めたり弱めたりする働き」を持ち、前頭前野の“司令塔の司令塔”とされています。OFC・ACC・DLPFC はそれぞれ違う機能を持つネットワークです。
OFC:情動・報酬・不安 ACC:エラー検出・葛藤 DLPFC:実行機能・抑制・ワーキングメモリ
MD 核は、これらのネットワークの「どれを優先的に使うか」を調整します。例:不安が強い状況 → OFC 系を強める ・認知制御が必要
→ DLPFC 系を強める つまり、状況に応じて前頭前野の“モード切り替え”を行うのが MD 核です。
視床MD核はこの機能があるため、淡蒼球内接を通過して視床に投射された行動計画に加算ポイント(ゲイン)を与えます。行動Aの興奮度が100点なら130点と30点ゲインを与え、大脳皮質(OFC ACC DLPFC)に返送します。これは、行為そのものよりも、「その行為を選択すべきだという信念や不安表象」にゲインを与えているからとされています。
第5節 出発点に戻ってきた行動計画
かくして行動計画は元の皮質(DLPFC・
DFC・
ACC・島皮質)に戻ってきました。しかし、行動内容は同じでありながら、不安・葛藤はより増強しています。線条体で過剰なドパミン投射(報酬予想誤差)、偏桃体や海馬から記憶や恐怖感をベースにしたグルタミン酸投射(興奮性)、また視床では拡大ゲインを得て、(OFC ACC DLPFC)「赤文字部位」がより刺激されています。これの意味するところは、行動計画→行動実行→一次的安心感→再び不安感→行動計画→行動実行(強迫行為)・・・・と無限ループに陥ってしまうことであり、強迫症が文字通り発症してしまうことです。
第6節 習慣経路の生成
そしてこの過程の中で、強迫症をより難治性にしてしまう事態が発生します。習慣回路の生成です。この回路は、皮質(運動前野・感覚運動野)→ 背外側線条体 → 淡蒼球
→ 視床 → 皮質 というループで、運動習慣・行動習慣の自動化を担います。今までメインで取り扱っていたCSTC回路(尾状核経由)とは、尾状核→背側被殻で連絡する必要がありますが、単純な一本の直結線ではなく、中脳ドーパミン系や(線条体‐黒質‐線条体スパイラル)と呼ばれる段階的ループにより機能的結合によるものとされています。
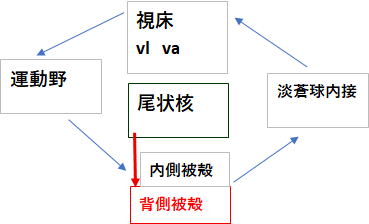
この習慣回路の形成は強迫症をより難治性にしてしまいます。
第一にネガティブ強化はポジティブ強化より強力で修正が効かなくなります。 ネガティブ強化とは、
不安・恐怖・不快感 これらが 行動後に減る ことで行動が強化される現象です。脳は「危険を避ける」学習を非常に優先するため、 不安が下がる経験は強烈な記憶として固定されます。そのため、 “不安が下がった”という経験は、報酬(快感)よりも強く行動を強化する という特徴があります。
第二に習慣回路が出来たことによって、不安・エラーを作るCSTC回路(ACC・ OFCが担当)と行動パターンを実行する回路(DLPFC担当)が分業化してしまい、二重の駆動力が同じ行動に向かう状態に陥って強迫症がより難治性になってしまいます。。つまり、強迫観念(OFC–尾状核) × 強迫行為(背側被殻) という二重構造が強迫症の本質なのです。
第7節 強迫症に対する調整系機能
ここまでの流れだけを見ると、「強迫的ループは必然的に暴走しそう」に見えますが、実際には脳にはそれを抑える調整系が複数あります。
①セロトニンによる調整作用
セロトニンは前頭前野に作用して抗不安作用をもち、また線条体に広く作用して D1/D2両方の可塑性にブレ ーキ的に介入 し「やりすぎ」を防ぐ方向に働きます。
②ハイパー直接路の作動
ハイパー直接路の機能は、「即時に全体へブレーキをかける回路」で、強迫行為が続く場合に作動します。
③背外側前頭前野(DLPFC)
のトップダウン制御
トップダウン制御とは、「考えて行動をコントロールする力」のことです。「やりたい」「気になる」
という衝動に対して 「今はやらなくていい」とブレーキをかけ、違和感や不安感に対して「別のことに意識を向 ける」よう注意の切り替えを行います。最も強力なのは、「仕事や学校に遅れる→社会から
ドロップアウトする→生存が脅かされる」という思考を持ち出し、ネガティブ(不利益)と強迫行為を続けるメリット(不安低下)」の価値比較を行うことです。これにより、 ネガティブ vs ネガティブ の価値比較が 行われ、 “より大きな不利益を避ける”方向に行動が切り替わります。
以上のような調整系を作動させても強迫行為が続くのは、「
抑制はしているが、“十分に勝てていない」つまり、「自助努力での自然治癒は難しい」ということであり、一日も早く受診し治療(薬物治療と認知行動療法)を開始する必要があります。
第2章 ピッタリ系強迫症の発症原理
第1節 思春期の感覚統合
思春期の脳は発達特性上、「違和感・エラーを検出する回路」が 敏感になります。
これは第一に、環境への適応精度を上げるためです。思春期は社会へ積極的に参加して新しい人間関係を築く時期なので、微細な違い・ズレを検出する能力が高い方が適応しやすい つまり社会的サバイバルに有利となります。
第二に学習効率を上げる(誤差駆動学習)ためです。脳は基本的に「予測と現実のズレ(誤差)」で学習するとめ、誤差(=違和感)に敏感な方が学習が進みます。
このようにして、社会人としてのルールを学び、知識の基礎を習得した後に、「それを抑える・流す回路」が
後から発達してきます。つまり、思春期とは「ブレーキよりセンサーが先に強くなる時期」であるとされます。ピッタリ系強迫症は、感覚統合(特に左右差や“しっくりこなさ”の処理)が過敏になることが発症のベースにあるので、思春期に発症患者が多いのは一応理に適っていると思われます。
第2節 感覚統合とピッタリ系強迫症
感覚統合とは、一次感覚野(位置、強度、方向など)→二次感覚野(複数の感覚情報を統合)→.頭頂葉(身体位置、空間認知、左右の整合性、身体図式の統合
)→前頭前野 ( 感覚情報を「意味づけ」し、行動選択に結びつける)と複数の領域が協調して働くシステムです。
ところがピッタリ系強迫症は、感覚統合(特に左右差や“しっくりこなさ”の処理)が過敏になる、つまり「違和感・エラーを検出する回路」が
敏感になっているので、その情報が前頭前野の主要部位(OFC ACC DLPFC)に投射され、強迫症の基本パターンそのものとしてピッタリ系強迫症が始まります。
机の上の物の配置や食器の置き方にこだわる、ノートの升目にピッタリ収まらないと書けた気がしない、などは典型的なピッタリ系強迫症の症状といえます。
しかしピッタリ系の場合、典型的なパターン以外にも、特定順序・ 特定回数
・ 特定運動 にこだわるなど解釈が困難なケースも存在します。そこにはピッタリ系強迫症独自のエラー回路の存在が推察され、そのうちの一つがPVニューロン障害と思われます。
第3節 感覚統合とピッタリ系強迫症
これは誰しも経験のあることかもしれませんが、何か不安があると自然と「繰り返し行動」を行うことがあり、いつしかそれが癖になったりします。つまり:不安定↓反復運動↓神経系が安定 という構造です。
おそらくPVニューロンの障害もこれと同じ作用原理で、ピッタリ的強迫行為が発生してきます。
PVニューロンはgaba投射性の介在ニューロンです。皮質からの行動計画と線条体の間に介在して、行動計画を線条体に伝える働きをしています。ところでこのPVニューロンはCa²過剰に非常に弱く、Ca²⁺が過剰になると、発火リズムが乱れる・γ同期が崩れる・MSNへの抑制が弱くなるなど、 行動選択のゲートが壊れてしまいます。
繰り返すと、思春期は「違和感・エラーを検出する回路」が過敏になる時期です。もともと不安回路が敏感な人は、この時期に“生き残るためにもっと正確でいなければ”という感覚が強まり、内側の違和感が脳の警報システムを刺激します。しっくりこない 左右が揃わない 感覚がズレている 何かが“間違っている”気がする。これらは脳にとって「危険の予兆」として扱われやすく、
社会的な負荷が増える時期にその感覚が強まるのは自然な流れです。ところが、この違和感は脳に急性ストレス(瞬間的ストレス)をもたらします。そして、この急性ストレスは、交感神経・グルココルチコイド系を一気に活性化し、細胞内カルシウム濃度を急上昇させます。
結論として 「瞬間的なストレス → 一過性のカルシウム過剰 → 線条体に隣接するパルブアルブミン(PV)ニューロンの瞬間的機能障害」と結びつき、行動選択のゲートを壊してしまいます。図()でDLPFCが担う具体性のある行動計画(違和感を修正する)が反映されないと、違和感のみが線条体でマッチングされてしまい、カーナビのない車のように道に迷ってしまいます。多分その時には、慣れ親しんだ道に戻るのが結局正解だったりします。線条体は「最適」ではなく「最も強化済みpolicyを選ぶ」とされています。つまり、線条体は必ずしも、最も合理的 ・ 最も正確
な行動を選ばないで、むしろ、「以前少し違和感が減った行動」を優先します。
幼児期には、揺れる・ 触る・並べる・
回す、など
同じ動きを繰り返す反復感覚運動行動が非常に多い。ただし、これらの反復行動は単なる未熟さではなく、覚醒調整 不安低減
感覚統合
予測可能性確保
の役割があるとされています。そして、これら発達初期に形成された、感覚運動パターン・
早期習慣回路・
自己鎮静行動 が、ストレスや統合不全時にピッタリ行動という形で表れてくると考えられています。
つまり、左右対称・ 感覚的一致・ リズム・完全性・ 身体感覚補正 など、発達初期に強く強化され、感覚運動的に安定化し 認知負荷が低く、自己調整に有効だった 行動policyが、脳の不安定時にピッタリ系行動として顕在化するというイメージです。