![]()
![]()
複眼は節足動物の昆虫類と甲殻類に限られていますが、なぜこのような眼をもつようになったのでしょうか? これは彼らの特徴である外骨格という堅い皮膚をもっている事が大きく関係しています。複眼はこの皮膚構造を基本にできあがっているのです。正確に言えば、幼虫の上皮細胞から構成される眼原基から複眼が作られますが、これは皮膚と相同の器官といえるのです。同じような器官としては、毛の動きで空気振動や触覚を感ずる感覚毛や皮膚の歪みや振動をキャッチする弦音器官と呼ばれる感覚器があります。特に弦音器官は皮膚内部に神経細胞を持ち、複眼と非常に似た構造をもっています。
皮膚の構造については体色の項で後日紹介しますが、昆虫の表皮は複数の真皮細胞(色素細胞)により小さな区画(パッチ)に区切られ、真皮細胞から分泌されるクチクラにより堅い表皮(外原表皮)と比較的柔らかい表皮(内原表皮)が形成されます。これが集まって外骨格を形成しているのです。また真皮細胞の下には基底膜があり体腔と隔てられています。
【個眼の構造】
個眼は円錐晶体の形態により、いくつかの型に分かれます。通常は“正晶子体眼”と言われチョウ、トンボ、バッタ、ハチなどの複眼が代表的例で、通常の円錐晶体を持つ眼です。晶子体細胞はあるものの、全く円錐晶体が形成されない眼を“無晶子体眼”といい、テントウムシ、カミキリムシなどがこの型に属します。また堅い晶子体はなく、透明な半液状物で構成されている眼を“擬晶子体眼”といい、ハエなどがこの型に属しています。更に晶子体細胞が円錐晶体を形成せず、代わりに角膜レンズの内側が伸長して角膜晶子体となり、機能的に円錐晶体の役割を果たしているタイプもあります。これを“外晶子体眼”といい、ホタルなどがこれに属します。
個眼が集まった複眼には大きく2つの種類があります。連立像眼と重複像眼です。これらを説明する前に、まず複眼を構成する個眼の構造から紹介しましょう。
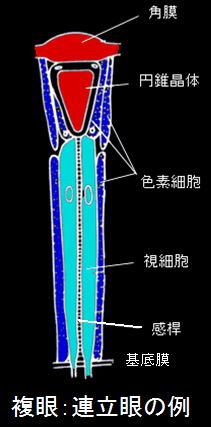 |
既に説明しましたが、個眼は、一番外側にある角膜、角膜の下にある円錐晶体、円錐晶体の下部を取り囲む視細胞、体腔との仕切りである基底膜から構成されています。視細胞の最も下部の基底膜からは神経軸索が出ており、1次視覚中枢である視葉につながっています。また複眼には全体の光量調節を行う虹彩はありません。必要に応じて個眼の円錐晶体や視細胞を囲んでいる色素細胞にある黒い色素顆粒が移動して光量を調整します。
個眼の大きさは通状、20〜40μですが、中には100μもあるものがいます。個眼の表面をなす角膜は単なる膜ではなく、特殊化したレンズです。この表面は凸レンズ状、平坦やデコボコ、またひげがたくさん生えていたりと様々です。この表面形状は種により様々ですが、生きた化石、カブトガニでは個眼間の区画がなく平坦な表面をしています。ザリガニも平坦な表面をしていますが、同様にカブトムシなど夜行性の種では平・凸レンズ状、昼行性のハナムグリでは両凸レンズ状をしています。また角膜の屈折率も種により異なり、コガネムシでは一様ですが、中心部の屈折率が周囲よりも高い種もいます。またアブでは表面近くに屈折率の異なる層が交互に重なり干渉フィルターとなって、干渉効果により眼に色の縞模様を出す原因になっています。角膜の外面形状には、六角形と正方形の2種類しかありません。表皮の限られた面積にたくさん個眼を詰め込むためには六角形が最も稠密充填できることが良く知られていますが、これと関係しているようです。ちなみに個眼数の多いトンボでは六角形、アブでは通常、中央が六角形で周辺部は四角形になっています。一方、ザリガニの中央部の個眼の形状は四角です。伊勢エビも四角い形状をしていますが、カニは少し異なり、六角形をしています。シャコは種により異なるようです。またショウジョウバエには個眼間にひげ(剛毛)が生えており、刺のようになっています。一般的には、六角形の個眼は明所で暮らす連立像眼をもつ動物に多く見られるといわれています。
さらにチョウやガなど鱗翅目やある種の脈翅目の昆虫ではこの角膜の表面に、反射防止機能をもつ事が確認されています。ガでは、角膜表面に高さや間隔が200μ程度の小さな凹凸をもっていますが、これで反射光を低減し、天敵から目立ちにくいようにしているようです。
角膜の下の円錐晶体はレンズの働きをしていますが、円錐晶体の下には光を受けるための特殊な構造が作られ、この部分を“感稈”と呼びます。またこの感桿部を囲むように、円錐晶体の下部に複数の視細胞が存在しています。感稈部ではこれらの視細胞から光を感ずる小さな毛(微絨毛)が多数突出しています。この微絨毛部には光を受容する視物質が多量に含まれており、この多数突出している部分を感稈分体といいます。この微絨毛は大部分が細胞膜で構成されていますが、細胞膜はリン脂質が多く含まれるため、結果的にこの感稈部は周囲よりも屈折率が高くなり、光ファイバーのような屈折率分布をもつ構造になっています。
一方、色素細胞は色素顆粒を含み、明暗順応によって変形したり、中の色素顆粒が移動する等、光量を制御する役割を果たしています。色素細胞には2種類あります。1つは虹彩色素細胞(一次色素細胞)といわれ2個の細胞からなります。他は網膜色素細胞(二次色素細胞)と言われ、昆虫の種類により変化しますが、通常は12個の細胞で構成されます。これら色素細胞の一方の端部は角膜レンズに密着しています。虹彩色素細胞は晶子体を環状に包みこみ、網膜色素細胞は隣接する個眼間に存在して視細胞を取り囲み、下端は基底膜まで達する事があります。これら色素細胞内の色素顆粒は0.5μ程度の大きさですが、夜行性昆虫に見られる重複像眼ではこの色素顆粒の移動が見られます。
【視細胞】
昆虫では個眼の視細胞は通常は8個、ミツバチでは9個から構成されます。但し例外もあり、中には6個や16個ももつ種も存在しています。これら視細胞は神経細胞の樹枝状突起に相当しています。また視細胞の受光部は、細胞の形質膜が規則的な6角形をした微小管状をして個眼の中心軸方向に突出しています。
これらの視細胞から構成される感稈部は感稈分体が各々隣接して1つに集まったものとばらばらになっているものがあります。ミツバチ、セミ、アゲハチョウでは一体になっていますが、ハエやアメンボなどでは感桿分体は隣接せず、ばらばらになっています。
また感稈の周囲は色素で取り囲まれています。この色素の色は昆虫の種類により異なり、ミツバチには色素はみられませんが、アゲハ蝶では赤や黄色の色素、モンシロチョウでは赤と暗赤色の色素、シジミチョウでは紫色の色素で囲まれています。個眼で受光した光は視細胞で受光される前にこの色素による色フィルターを通るために、視細胞の分光感度はこの影響を受ける事になります。特にチョウなど感桿分体がばらばらになっているケースでは、感桿部の上部(角膜側)に存在する視細胞では、光の通過距離が短いために、この色フィルターの影響はあまり受けませんが、感桿部の下部(脳側)に存在する視細胞はこの色フィルターの影響を強く受ける事になります。この様に、色フィルターと異なる視物質を組み合わせる事で昆虫は多様な視細胞を持つようになりました。ちなみにアゲハチョウでは5種類、モンシロチョウでは4種類の視物質があり、色フィルターと組み合わさって多様な色を受容しています(なおミツバチの視物質の数は3種類です)。また紫外線感知能力がアリで初めて発見されましたが、紫外線を受容する視細胞をもつ事は昆虫では例外的な事ではなく、昆虫の色覚に共通の特徴です。
ちなみにミツバチでは紫外線、青色、緑色の3種類を感ずる視細胞が、アゲハチョウでは紫外線、紫色、青色、緑色、赤色、広帯域の6つの波長域を感ずる視細胞が確認されています。赤い花はミツバチ(ハナバチ・スズメバチやアリも同様(注アリはハチの仲間です)には見えず、アゲハチョウは見える事になります。またモンシロチョウにも、紫外、紫、青、黄緑、赤、暗赤の6色の視細胞がありますがモンシロチョウは赤色が見えません。このように一部のチョウを除き、多くの昆虫ではミツバチ同様、長波長の赤色を色としては見えません。ハエも赤い色は見えません。
ナミアゲハでは紫受容細胞を、モンシロチョウでは赤や暗赤受容細胞を吸蜜行動時に色を見るのには使っていないと考えられており、色感覚はアゲハ蝶では紫外、青、緑、赤の4色と考えられています。なお、タテハチョウ科のチョウでは赤い色の受容器をもたないチョウが多く見つかっています。またモンシロチョウでは角膜側のR1,2視細胞は雄雌で異なった組み合わせを持っています。雄の場合420nmの光で蛍光を発する個眼を持っており、雌では420nmの紫を感ずる視細胞が、雄では420nmの光がカットされた400nmと450nm前後にピークをもつ2山の光を特に感ずる様に変化しているのです。後述するように、モンシロチョウは雄が交尾相手の雌を探す際に雌の翅の紫外線反射を目印にしています。従ってこの目的を果たすために、特に雄の眼は短波長側の光の弁別能力を高めているのです。
なお、ミツバチでは短波長側の色ほど識別能力が向上し、夜行性昆虫では紫外部の光によく反応する傾向があります。昆虫は短波長の光が行動上重要なのかもしれません。
さらに、ほとんどのチョウには、個眼の基底部にタペータムがあり、吸収されなかった光を再度個眼に戻す働きをしていますが、このタペータムは毛細気管により光を反射させる構造になっています。なおアゲハチョウ科のチョウはタペータムを持っていません。アゲハチョウは他のチョウと比較し、視物質の数が多くなっており、これと光の利用効率と関係があるのかもしれません。
花色と訪花蝶の関係を調べた結果も多くありますが、カラスアゲハが断トツで赤い花を最も良く訪問し、アカタテハは白や緑色の花に、モンシロチョウやベニシジミ、ツマグロキチョウでは黄色の花を良く訪花する傾向が強い事が判明しています。但し、雌雄でも色の好みに差があるようです。
また、ミツバチやアゲハチョウでは視力の測定が行われています。アゲハチョウの方がミツバチよりも視力が高く、視力限界まで色が見えているようです。なおミツバチでは形を見る能力も良く調べられており、その結果では、左右の対称性やレイアウト情報なども良く認識できるようです。三角、四角や丸、また点や線などの区別もできる事が確認されています。
これらの事から昆虫は形の性質1つ1つを見ているだけでなく、視野全体の中でどのように配置されているかという事も認識し、この能力で、視覚的な地図を情報として利用している可能性が示唆されています。なおトンボのアキアカネは紫外線を含む4色が見え、ゴキブリは紫外線と緑色しかみえません。なお1つの視細胞が1つの色を感受するとは限らないようです。ミツバチでは1つの視細胞は1色の色しか感受しませんが、他の昆虫では2種類の色を感受するものもあり、複雑な構成をしています。なお、ミツバチでは中間色は識別できず、青、青緑、黄色の3色のみしか識別できないようです。アゲハ蝶についても中間色は識別できないと推定されています。
なお視細胞の微絨毛は非常に規則正しく配列しています。これは既に述べたように、昆虫が光の偏光成分を検知できる事と関係しています。視物質の配列に方向性がある事で、光の波が変化する方向の成分が分かるのです(太陽の光は大気層で屈折されるときに直線偏光性が出ます。観測者の頭上(天頂)と太陽を結ぶ線に垂直な偏光成分が多くなります。これは地球規模の現象ですが、これを動物は検出しているのです)。このような偏光を検知する能力はアリやハチ、バッタ、ハエ、チョウなどで確認されていますが、ミツバチやアリではこのような偏光を検知するのは紫外線を感知する視細胞がその役割を負っています。ミツバチでは複眼の背側(頭部の単眼側)に、ハエでも複眼の背側辺縁部に偏光を検出する領域があり、この部分の1つ1つの個眼の感桿微絨毛が1方向の偏光を検出するように並んでいます。このように昆虫では偏光を検出する個眼を複眼の特定の場所に配置する事が良く行われています。
この他に、夜行性のアリでは月の光をコンパスに用いる種も確認されており、セイヨウミツバチでは地磁気の利用も行われています。なおザリガニなどの甲殻類も偏光の検出ができます。夜行性のフンコロガシでは月による夜空の偏光パターンを見て帰巣方向を定めているといわれています。
このように個眼が集まって複眼が構成されていますが、複眼は主に昼行性の種に多くみられる連立像眼と、夜行性の種に多く見られる重複像眼に大きく2つに分けられます。