![]()
![]()
前回、鼻の構造について紹介しました。人間には約500万個、イヌでは2億個、ハト600万個、ウシガエルで約800万個、イモリで80万個の嗅細胞があるといわれています。これらの嗅細胞には1細胞当たり1つのにおい受容体しかありません。におい分子をこの受容体がつかまえると、受容体の分子構造が変化し、これが受容体と結合しているG蛋白質というものに伝わります。これが匂い検知の最初の変化で、最終的にはG蛋白質の働きを介して、イオンチャネルが制御され、嗅細胞の膜電位が変化(“-”から“+”)します。この電位変化が嗅細胞から主嗅球に、また鋤鼻器の嗅細胞で検知された情報は嗅球の後方にある副嗅球に送られます。嗅球にある個々の糸球はただ1種類のにおい受容体情報のみを受け取るため、脳では活性化された糸球体の組み合わせパターンからにおいを判断しています。従って、嗅細胞の数ではなく、におい受容体の種類数により動物の嗅覚の性能が決まる事になります。
なお、嗅上皮の嗅細胞からは多数の軸索が出ており、はじめはランダムに嗅球につながっていますが、匂いを嗅いで神経が活動することで特定の糸球体との関係が強化され接続が確立してゆくようです。従って、訓練を積む事で嗅覚は鋭敏になってゆく事になります。また、地球上には数十万種以上のにおい物質があり、この中で、人間は約2~4万種のにおいをかぎ分けているといわれています。それではこのにおい受容体にはどのような種類があるのでしょうか?
匂いを検出する嗅覚受容体には以下のようにいくつかの種類がある事が判明しています。
① OR:odorant receptor
② TAAR:trace amine-associated receptor
③ V1R:vomeronasal receptor type 1
④ V2R:vomeronasal receptor type 2
⑤ FPR:formyl peptide receptor
最初のOR受容体は、動物が匂い物質を検知する主な受容体ですが、V1R・V2Rはフェロモンを検知する受容体です。またTAARは生体アミン受容体です。また最後のFPRはフォルミルペプチド受容体で最も新しく見いだされた受容体です。ORやTAARは嗅上皮に、V1R・V2RやFPRは主に鋤鼻器に存在しています。またこれらの種類は動物種により異なります。OR受容体を最も多く持つのはマウスやカエルで1,000種以上(マウスで1,063種、カエル1,024種)あります。イヌも多く872種、ウシでは1152種、人間は388種保有しています。哺乳類以外では、魚類では50~100種程度、鳥類(ニワトリ)で300種、爬虫類(ミドリアノール)で116種となっています。一方、昆虫類では60~70種程度ですが、ミツバチは多く163種、またコクヌストモドキという昆虫では262種と例外的に多くもっています(なお、これらの数は文献により値が異なりますので大小の比較材料として考えてください)。
嗅覚器と情報伝達経路例:マウス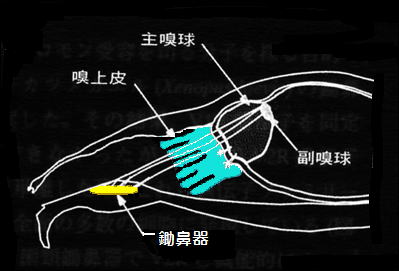 |
脊椎動物のOR受容体はナメクジウオにも存在する事から、嗅覚受容体の起源は脊索動物の共通祖先までたどる事ができると考えられています。またこのOR受容体は大きく2種類に分かれます。1つはクラスⅠと言われ、魚類のOR受容体に類似する水溶性の匂い物質を受容するタイプで、他はクラスⅡと言われるもので、陸棲動物に得意な受容体で空気中に揮発された匂いを受容するタイプです。クラスⅡは水棲から陸棲へと生活環境が変化するに従ってクラスⅠから進化したものと考えられています。陸棲動物は水棲動物よりもOR受容体の種類が多くなっているのはこのような理由によります。またOR受容体を形成するOR遺伝子を昆虫と脊椎動物で比較すると遺伝子配列は異なっており、これらは別の進化過程をたどったようです。また、ミツバチでは花のにおいを認識するとともに、社会性の維持に(各種の信号フェロモンを認識するのに)匂いを用いているため多くのOR受容体を活用していると思われます。さらに昆虫では、植物の出すにおい物質(昆虫により食害を受けているというシグナルや“みどりの香り”といわれる揮発性化合物等)も感知している事が分かってきました。
TAAR受容体はアミン系匂い物質の受容に特化した受容体ですが、魚類が100種程度と多い反面、他の動物では数種、多くても10数種と少なくなっています。魚類はアミノ酸の匂いに非常に敏感です(味覚も同様)。また、VR1は鼻腔の内腔側にあり、揮発性のフェロモンに応答し、VR2は基底側にありますが、不揮発性フェロモンを受容しているのではないかと推測されているようです。助鼻器をもつ動物はこれらの受容体を多くもつ事になりますが、マウスではVR1は187種、VR2は121種、カエルの場合、VR1は21種、VR2は249種もっていますが、人やイヌではVR1は数種、VR2はもっていません。魚類(ゼブラフィッシュ)ではV1Rは2種、VR2は44種もっている事が確認されています。魚類は鋤鼻器をもちませんが、嗅上皮で匂い物質とフェロモンの両方を受容していると考えられています。なお、人間のフェロモンについてはまだその存在についてはっきりとした結論は出ていないようです。
また、サケなどは産卵のために母川に戻ってきますが、この母川の判断は匂いにもとずいていると言われています。この点について研究した結果では、魚類の匂い物質としては、①アミノ酸とその関連物質、②無機塩類、③胆汁酸、④ステロイド類、⑤他、等があり、①と②以外の匂い物質は、ほとんどが同種の魚から分泌・輩出されたものに由来する(従って同種の匂いでなければ敵か餌の匂い)事から、母川の匂いは①のアミノ酸、ではないかと考えられています。
なお、最後のFPRは細菌由来のペプチド分子を受容し、感染症の有無など、他個体の健康状態を認識する役割を担っている可能性が指摘されています。人間・イヌ・マウスなどはこの受容体を数種類もっていますが、ゼブラフィッシュ(魚類)は保有していません。
以上のように、動物では哺乳類(霊長類を除く)が最も嗅覚が発達し、脊椎動物では魚類が比較的嗅覚に乏しいようです。但し、魚類の場合、味覚も情報源として使用しており、特に、ナマズやコイなど泥水の中で棲息し視覚情報に頼れない種では、体表に広く味蕾が分布しており、味の濃度分布から餌対象の位置まで計測できるようです。またウナギなども嗅覚が発達していると言われています。鳥類の場合、視覚が発達しているので有名ですが、嗅覚が比較的発達しているのはコンドルなどの腐肉食性の鳥類、アホウドリなどの海鳥類、キウィなどの地上性の鳥類や地上に巣を作る種といわれています。また夜行性の種の方が、昼行性の種よりも嗅球が大きいといわれており、夜行性に移行した歴史をもつ哺乳類で嗅覚が発達している事ともよく対応しているようです。