第1章 チック症
次頁 総合案内
第4節 病態論
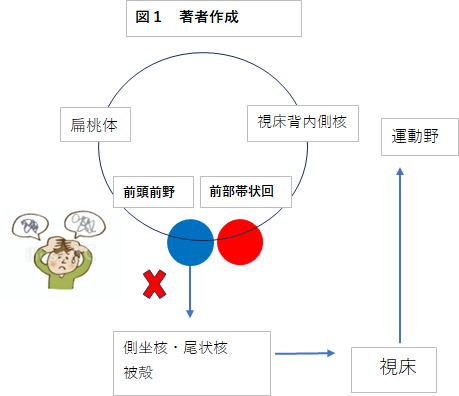
瞬きをトリガーとした、「ドパミンと行動の運動モデルの内部化」が完成しないとどうなるのでしょうか。そして、それがどうチック症に結びつくのか。舞台は前頭前野に移ります。前頭前野はヤコブレフ回路に属して、行動計画をプランニンングする部位でした。
(前頭前野のプランニング不調)
上図をもとに説明すると、前頭前野で計画された行動プランはCSTC回路に入り、運動野経由で前頭前野に戻ってきます。ところがチック症患者ではおそらく少なくとも二つ問題があり、この経路による行動プランは中々遂行されません。
一つは「行動とドパミンの連携」に関わる内部モデルの形成が一部未完成になっていることです。
そして第2にヒスタミンH3受容体です。これについては、
※chat-gptで調べた「前頭葉とヒスタミンH3受容体の関係」の文章を紹介します。
「ヒスタミンH3受容体は、脳内でヒスタミンの神経伝達物質として機能し、覚醒状態の維持や認知機能に関わると考えられています . 前頭葉のヒスタミンH3受容体と作業記憶が関係していることが発見されています」
また、このヒスタミン関連遺伝子とチック症の関係については、
※ 博士論文
論文題目 トゥレット症候群の De
novo 変異の探索 江里口 陽介
「多数のトゥレット症候群患者のいる家系からは、ヒスタミン合成の
律速酵素である L-ヒスチジン脱炭酸酵素をコードする、HDC が候補遺伝子とし
て報告された。HDC ノックアウトマウスには、チックに類似した常同運動
がみられるため、トゥレット症候群のモデル動物として研究されている」
これらことから、ヒスタミンやヒスタミンH3受容体の機能異常が前頭前野の行動プランニング遂行に何らかのマイナス要因を与えていると考えられます。
以上の内容から、前頭前野の行動プランはうまく遂行されません。上図には赤×が記載され、当人はおもわず頭を抱え込んでいます。ちなみに、ヒスタミンは痒みの感覚に関係する神経伝達物質で、チック(トゥレット症)患者のイライラ感にも関係あるとされています。
そこでこの窮地を救う脳内システムこそが、チック(トゥレット)症の症状そのものに直結することになるのです。
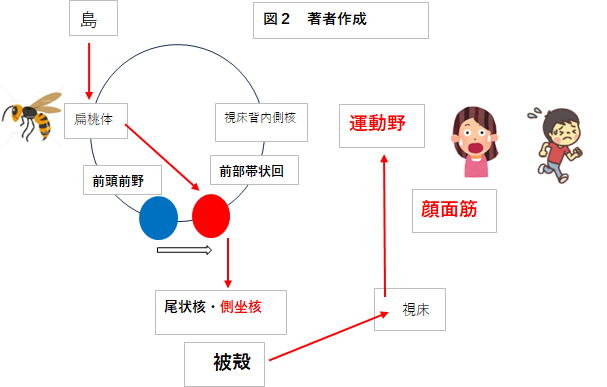
症状の多さは、偏桃体が示すバーチャルな対象がスズメバチの他、個人の感情への応答性に対応していくらでも
増やすことができます。例えば、ヘビ・金塊・食事・異性(性的なもの)などです。要は前部帯状回が十分に賦活
すれば良いのです。そして結果としてのリアクションがチック(トゥレット症)の症状になる。これが私が考える
チック症の根本的なメカニズムです。
この仮説を後押ししてくれるような研究報告もなされています。
※チック症の発現機序を紐解く-光イメージング法を用いたアプローチ- kaken
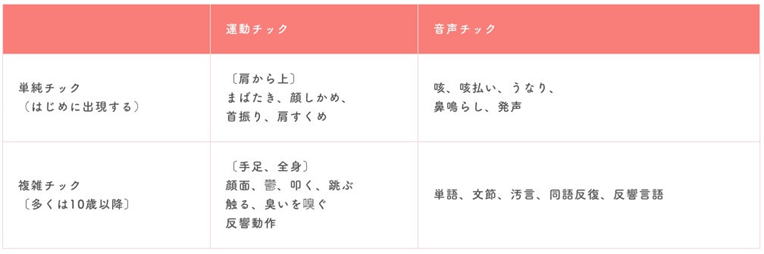
チックとは、反復性の動作を示す運動チックと奇声を発する音声チックから成るが、その病態に関しては明らかでない。線条体に
おける異常興奮がチックの本態であるという仮説のもと、マウス線条体へのGABA受容体拮抗薬注入により、一過性の筋収縮を示す
チック症状を呈するモデルマウスを作製することに成功した。さらに、c-Fos蛋白の免疫染色を行うことで、症状発現時の活性化
脳部位を検討した結果、一次運動野に加え、扁桃体、帯状皮質、島皮質といった情動機能に関与する辺縁系脳部位の活性化を観察
した。このことから、チックは運動のみならず情動に関連する脳領域のネットワーク異常に起因する疾患であることが明ら
かとなった。
※トゥレット障害の発症メカニズム解明に新展開 -音声チック症状を呈する霊長類モデルを開発- 京都大学
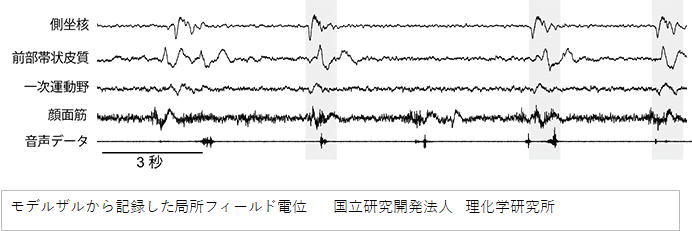
韓国と日本の複数の研究機関からなる本研究チームは、側坐核と呼ばれる脳部位の活動を興奮状態にすることにより音声チックを
再現できるモデルザルの作出に世界で初めて成功しました。このモデルザルの脳活動をPETで調べたところ、発声に関わることが知
られている前部帯状皮質という部位で脳活動が過剰に亢進していることを見出しました。さらに、側坐核、前部帯状皮質及び発声
運動に関わる一次運動野(特に口腔顔面領域)の各部位の神経活動を電位測定により調べたところ、これらの部位の神経活動が
同期することによって音声チックの症状が発現するという脳のメカニズムが明らかになりました。 (中略) 次に、このモデルザ
ルのどの脳部位に異常が生じているのかをつきとめるため、放射線医学総合研究所の研究チームが協力して、脳全体の活動が計測
可能なPET(陽電子放出断層撮影)を用いてモデルザルの脳活動を計測しました。その結果、前部帯状皮質や扁桃(へんとう)体な
ど、発声や情動と深く関わる部位で脳活動が過剰に亢進していることを見出しました。
両研究とも、運動と情動に連結するネットワークがチック発症の原因とみていることが共通しています。
運動の駆動力はドパミンで、これがチック症に関連しているとの予想は一般的なものです。一方、情動はその利用
回路がヤコブレフ回路で、前頭前野と前部帯状回が行動プランニングを行っています。
ドパミンに対する行動量の目安が作成されないと、この影響は大脳基底核から大脳皮質へ連鎖します。つまり、
前頭前野で行動計画が作成できなくなってしまうのです。
そこでバーチャルなスズメバチが登場して、行動プラン作成の主体を前頭前野から強制的に前部帯状回に切り替
えます。結果、その行動命令はチック症になって表れるのではないか。これが私の提唱する、チックの2段階原因
仮説です。