統合失調症のドパミン仮説発見が偶然の産物だったように、グルタミン酸仮説も似たようなストーリーを持っています。1970年代のはじ め米国ワシントン D.C.において、抗精神病薬に抵抗性の症状をもつ初発の統合失調症患者の発症数が、突然通常の 3倍になる という事態が生じ調査を行ったところ、麻酔薬のフェンサイクリジン(PCP)が乱用された結果と分かったことに始まります。その後、PCPが非競合的に NMDA受容体を遮断する ことが分かり、統合失調症の グルタミン酸仮説(NMDA 受容体仮説)が提唱されるようになったのです。この仮説のドパミン仮説と比しての最大の注目点は、PCP乱用の症状が、陽性症状・陰性症状・認知機能障害を全て併存しており、専門の精神科医でも統合失調症と区別できないほどの臨床像を示したことです。
(グルタミン酸の役割と各種受容体 )
脳は興奮性と抑制性のニューロンから構成される神経ネットワークの集まりからできています。ちなみに、興奮性ニューロンとは、その生産する神経伝達物質がシナプス後細胞を脱分極させ、活動電位の発火を起こすことのできる神経細胞です。今回の仮説では、興奮性ニューロンがグルタミン酸細胞、神経伝達物質がグルタミン酸、その受容体がNMDA受容体(他にAMPA受容体)です。重要なことは、グルタミン酸が、脳内の神経活動の基になる神経伝達物質であるということです。 NMDA受容体は、イオンチャネル型受容体であり、後シナプスにスパイン構造を作ります。四量体構造を持ち、この開閉でNa+など正イオン流入の調整を行っています。
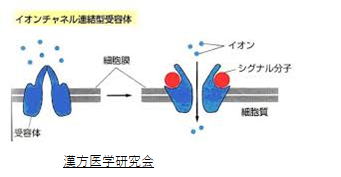
(AMPA 受容体)
グルタミン酸のもう一つの重要な受容体はAMPA 受容体です。AMPA 受容体はNMDA
受容体と共発現
しています(下図参照)
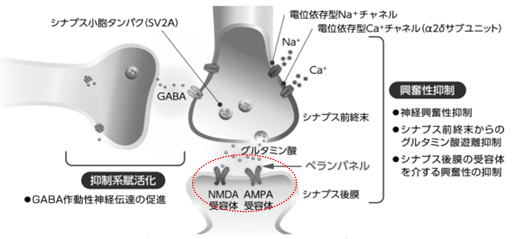
それではなぜ共発現しているのかについて、著者が理解(想像も含めて)しているのは次の2点です。
① 第一に、両者は取り扱う情報の分担をしている。 AMPA 受容体はグルタミン酸によってもたら
される興奮性神経伝達の大部分を担っています。これは、朝、起きてから寝るまでの意識せずとも
行う行動の全てに関与している部分です。一方で、判断(または思考)を要する行動もあり、
これに関する部分こそがNMDA受容体の担当するところです。それゆえNMDA受容体の機能不全
は統合失調症の認知機能障害に結びつきます。
② 第二に、AMPA 受容体とNMDA 受容体の連携が記憶素子(長期増強)形成に関与しています。
ここの部分は、第一章の「神経伝達の仕組み」で述べましたが、脳内の情報伝達で神経伝達
物質を使用する第一の目的は、記憶素子を形成するためためでした。
NMDA 受容体はカルシウムイオン(Ca2+)
透過性を持つので,高頻度刺激時の NMDA 受容体
活性化により、 Ca2+がシナプス後細胞内に流入します。すると、シナプス後部に流入した
Ca2+が引き金となって、タンパク
質リン酸化酵素などが活性化され、結果としてシナ プス後膜
上の AMPA 受容体の数の増加や,AMPA 受容体のイオン透過性の上昇が生じます。
その後もグル
タミン酸に対するシナプス応答が大きいままで維持されるようになります。神経生理学ではこれを
「神経可塑性」と呼んでいます。
可塑性とは弾性に相対する用語で、新たに形成された状態が維持されることです。ここでは、
後シナプスのAMPA 受容体の数の増加(シナプス体積の増大)が続くことを指します。結果、神経
可塑性はシナプス伝達の効率変化(記憶)をもたらします。
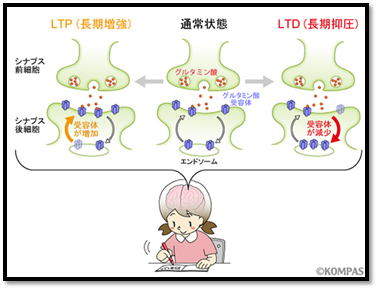
(NMDA受容体)
NMDA受容体は、不思議な受容体です。上述した通り、イオンチャネル型受容体で四量体構造をしています。
ところが、プラスイオンを透過する条件が、厳密に管理されているのです。他のイオンチャネル型受容体のように、シナプス内の神経伝達物質が受容体に結合すると、チャネルが開きプラスイオンを通過(透過)させるような単純構造ではないのです。NMDA受容体がプラスイオンを通過させる条件は、
①NMDA受容体にグリシンとDセリンが結合する
②グルタミン酸が結合する
③その上でチャネル内で蓋をしていたMG+が離れプラスイオンが流入する
という複雑な流れを有しています。
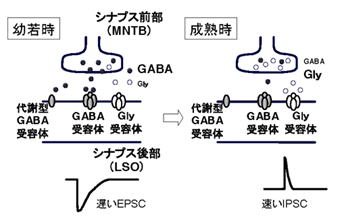
①のNMDA受容体にグリシンとDセリンが結合することが、NMDA受容体が働く必要条件になっていることについては、グリシンとDセリンが「NMDA受容体の働きを促進するアゴニスト」と従来、説明されていますが、著者の見立ては「γオシレーションに組み込まれているシステムの一端」とみています。
これ について、簡単に一部を説明します。
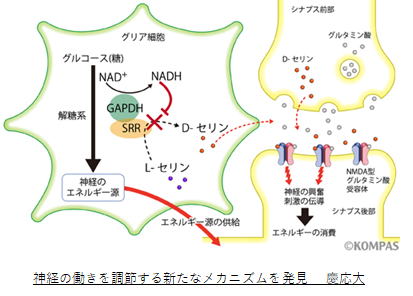
①1 まず、Dセリンはアストロサイト解糖系(エネルギーを作る)の副産物です。注目すべきは、
Dセリンの生産が解糖系の働きに反比例することです。解糖系の能力が限界に近づくとDセリンの
生成量は減少します。逆に、Dセリンがあるということは、解糖系がうまく回転しているということ
で、Dセリンの存在は、ATPが神経細胞に供給されることを保証するものです。
①2 それではグリシンのNMDA受容体への結合は何を意味するのでしょうか。γオシレーション
はNMDA受容体とGABA受容体の共同作業です。そのためには、神経伝達物質のグルタミン酸と
GABAが必要なのですが、GABAとグリシンは前シナプスの小胞に混載されシナプス間隙に共放出
されます。・・・・・・これから先はかなり複雑になりますので、(GABAー認知機能障害)の節
で改めて説明します。
※ 下図の二つの事実は最近(今世紀)解明された事実です。
※ γオシレーションについては、(GABAー認知機能障害)の節で説明します。
(NMDA受容体機能不全の原因)
統合失調症のグルタミン酸仮説は、NMDA受容体の機能不全から始まります。考えられる原因を下に
まとめました。簡単に説明すると
1 スパイン構造や細胞接着などのイオンチャネル構造形成に関する遺伝子変異(多型)によるもの。
2 グリシン
、D
─セリンがNMDA 受容体
に結合しないとNMDA
受容体 は機能しない。
それゆえ、この物質の代謝・放出・回収に関する遺伝子変異はNMDA受容体機能不全と結びつ
く。 例えば、Lセリンから Dセリンへの変換酵素 である(SRR)・(DAOA)・(DAO)等々
3 キヌレン酸は慢性炎症にともなって増加する物質で、これがNMDA受容体阻害作用を持つとされ
ています。
統合失調症患者に想定される NMDA 受容体機能低下の原因
1 NMDA 受容体 の遺伝子変異による構造変化
2 NMDA 受容体 に結合する物質の減少 グリシン D ─セリン
3 NMDA 受容体 の働きを阻害する化学物質濃度の増加 NAAG Kynurenic acid (キヌレン酸)
(NMDA受容体機能不全は何をもたらすか)
NMDA受容体の機能不全が、次の現象を引き起こします。
A グルタミン酸の過剰放出(脳内グルタミン酸濃度の上昇)
B 神経伝達物質gabaの減少
まずAのグルタミン酸の過剰放出について説明します。
ここでポイントとなるのが、gaba神経細胞上にNMDA受容体が存在することの理解です。この知識が本当に盲点になります。これが分かると、グルタミン酸神経細胞上にgaba受容体が存在することも理解できます。下図参照。
両神経細胞は、各受容体を介してループ構造となっています。
そして、このループ構造の知識はγオシレーション理解の重要なkeyとなります。
Aに戻ります。 下図を参照して下さい。 特にグルタミン酸細胞とgaba細胞のループ構造に注目
① NMDA受容体が阻害(×印)されると、gaba神経細胞へのグルタミン酸の刺激がなくなり脱分極
(発火)がストップする
② そのため、gaba神経細胞に電流が流れず神経端末からgabaが放出されない
③ gabaの抑制が外れた(脱抑制)グルタミン酸神経細胞は過剰に発火(電流の流れ)し、これが軸索
を通じて次の神経細胞へと伝わっていく
④ 過剰な電流によって、シナプスから大量のグルタミン酸が放出される
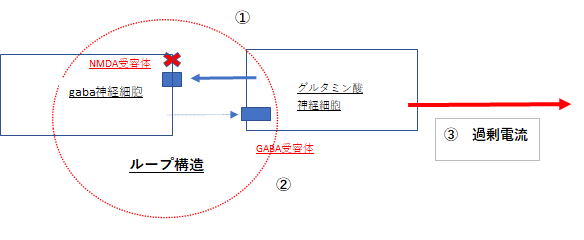
グルタミン酸の脳内濃度は、グルタミン酸神経細胞とgaba神経細胞の間のループ構造によって調整されます。これは極めて重要です。脳内のグルタミン濃度は高くても低くても、何らなの疾患と結びつきます。高ければ統合失調症の陽性症状、低ければパーキンソン病です。
グルタミン酸濃度はほどほどが良いのです。そして、そのほどほど加減を調節しているのが、上図のループ構造です。もし、そのループ構造がNMDA受容体の機能低下で変調をきたすとどうなるのか? これが、統合失調症のグルタミン酸仮説の本質です。
次にB
GABAはグルタミン酸から合成されます(下図参照 )。 その際、合成酵素であるグルタミン酸脱炭酸酵素(GAD)が必要なのですが、NMDA受容体が阻害されるとGABA神経細胞内GADのmRNA発現の減少(遺伝子発現の減少)をもたらし、結果的にGABAの生産量が減少します。
