第3節 勘合システム
(勘合システムとは)
まず最初に、私が提唱する拡大CSTC回路の概念、勘合システム(仮説)を説明します。
(概略)
行動計画は大脳の前頭葉で作成され線条体へ送付される、と同時に同じ内容が小脳・視床を通じて同じ部位の線条体へ送られる。
線条体は両データを照らし合わせて同じであれば行動命令として発する。違う箇所があれば、勘合誤差としてドパミン・アセチルコリンによる神経細胞の興奮性を調整し、またデータ情報(セルアセンブリ)を修正して、両情報を合致させる。合致された修正情報(行動計画)は脳神経細胞膜電位のup and down のup時(閾値超え)に行動命令として発せられる。しかし、両データの照らし合わせ作業がうまくいかない時には、前頭葉から送られた情報は未決処理され、被殻ループに乗せられ視床を通じて運動野へ送付される。これが、常同行動(強迫性障害の症状)となるが、無意味なものではない。
ちなみに、勘合とは日明貿易で使用された貿易の正当性を保証するシステムです。
※勘合貿易 日本と明がそれぞれ割賦を持ち交易現場の船上で合体させて使用した
上記概略を、以下に順を追って説明します。
(大脳皮質から線条体へ行動計画を発令)
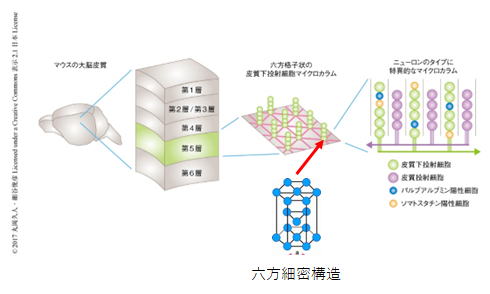
前頭葉は頭頂側頭連合野にある内部世界(操作される思考対象)を操って行動計画を作成します。この行動プランニングに際しては、大脳皮質にある情報単位(セルアセンブリ)を同期させる必要があり、大脳皮質全域で神経活動が活発になる。作成された行動プランは大脳皮質第5層から線条体へ同期情報として送付される。送付される行動プランは、どのよう な運動や行動を,どのような条件の下で,いつ起こ すかに関する情報です。ちなみに大脳皮質第5層は下表のようなカラム様構造をとっています。
※大脳皮質においてニューロンにより構築される格子構造 丸岡久人・細谷俊彦
(理化学研究所脳科学総合研究センター 局所神経回路研究チーム)
(ストリオソームとマトリックス)
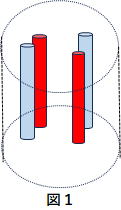
大脳皮質(C)から発せられた同期した情報を受けいれるのが、CSTC回路のSの部分の線条体です。この情報受け入れ機関(組織)の概要は次のとおりです(予想)。 まず個々の情報受領単位として図1のような構造をもつ神経構造が存在し、線条体の位置する箇所でストリオソームとかマトリックスと呼ばれます。基本は直径100~200ミクロンのコンパーメント構造の中に、直径3~18ミクロンの中型有棘神経細胞が複数入っています。この神経細胞は軸索末端から抑制物質gabaを放出するので抑制性の神経細胞です。 この中型有棘神経細胞は略してMSと表記されます。MSはD1とD2、2種類の神経細胞からなっています。
※青崎先生講義録 「線条体ニューロンの局所神経回路とその働き」
D1とD2は、ドーパミン受容体のサブタイプです。D1受容体を発現する中型有棘神経細胞は、直接路に関与しており、運動の促進に関わっているとされています。一方、D2受容体を発現する中型有棘神経細胞は、間接路に関与しており、運動の抑制に関わっているとされています。
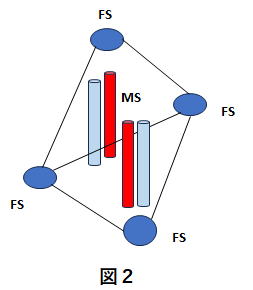
(MSとFS)
MSの周辺に介在神経細胞のFS(first
spiking interneuron)が配置されています。図2
介在ニューロンとは、投射系神経細胞(ここではMS)の近傍に位置して情報交換を行うニューロンのことです。FSはストリオソームやマトリックスのコンパーメント構造の中に入っているとの説がありますが、ここでは単にMSの周辺に介在ニューロン(FS)が配置されているということにします。
※青崎先生講義録 「線条体ニューロンの局所神経回路とその働き」
大脳皮質より同期してきた情報はFS回路網に入り、情報整理の後、線条体内での行動命令に必要な神経単位の組み合わせ(セルアセンブリ)をMS回路に描画(図3)します。 描かれた図はMSの位置(線条体のどこにあるか)と高さ(神経細胞の興奮のしやすさ=ドパミン量)を表しています。
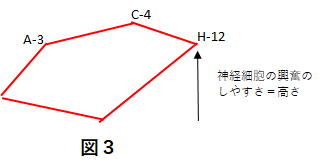
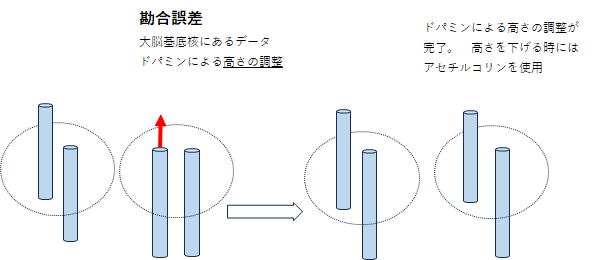
(ドパミンによる勘合誤差の修正)
図3で A C H などは線条体の位置を表していて、そこに大脳皮質のブロードマン領域のような機能が集約されていると想像します。外界の位置情報ならA、代名詞ならC、動詞ならHというような感じです。加えて、神経細胞の興奮のしやすさ(図3ではH-12と記している)がありこれにより、描画は3次元となります。 そしてこの高さの調節をしているのが、ドパミンとアセチルコリンです。ドパミンで高く、アセチルコリンで低くなります。
参考までに
他に重要な投射として黒質緻密部からのドーパミン
作動性入力がある.ドーパミン作動性の投射は大脳皮
質投射線維が線条体投射ニューロンの棘突起の先端に 興奮性シナプスを形成している近くにシナプスしてお り,線条体の投射ニューロンの活動性を変化させるこ
とにより線条体に入力する大脳皮質からの多様な情報 を取捨選択しながら出力される情報を調整していると 考えら れている
※大脳基底核回路の形態学的解析 藤山文乃
中でも コリン作動性ニューロンについての解析が進んで いる.中脳からのドーパミン作動性投射とコリン 作動性ニューロンとの相互作用が投射ニューロン の活動を調節する上で重要であり,基底核疾患で はこの相互作用がしばしば破綻する
※大脳基底核の機能;パーキンソン病との関連において 旭川医科大学 生理学第二講座 高草木 薫
(LTSによる描画修正)
さてここでCSTC回路について私見を述べます。「名は体を表す」と言いますが、CSTC回路には情報が一方向的に、皮質→線条体→視床→皮質に流れるものと思わせます。ところが、CSと同時にCTSの流れがあると考えると多くの事象と辻褄があいます。
CTSとは皮質→視床(小脳)→線条体です。図1から視床から線条体へ投射ニューロンが存在していることが分かります。
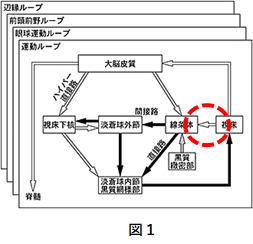
また視床と小脳は連結していて小脳には皮質と同じ内部モデルがあるとされています。CTSの流れは、皮質の行動命令に対し小脳の内部モデルで最適解を抽出して視床経由で線条体へ提示します。そして線条体でCSとCTSが合流します。CSでは線条体に3次元のセルアセンブリを描画しました。同時にCTSも線条体に3次元のセルアセンブリを描画するのです。描画される各点はストリオソームとかマトリックスとか呼ばれる中型有棘神経細胞(MS細胞)でコンパーメント構造の中に格納されています。
CTS経路による描画先は、CS回路と同じく、コンパーメント構造の中に格納されている神経細胞です。ここになぜMSはコンパーメント構造の中に格納されているかが分かります。つまり格納する構造がないと、CSが選ぶMSとCTSが選ぶMSとがワンペアとはならず、他のmSの中に埋没してしまうからです。
CSで流される情報と、CTSで流される情報。両者が線条体でお互いの情報を突き合わせる。このシステムは、日明貿易の勘合貿易に似ているので私は勘合システムと名付けました。以下、勘合システムの名を使います。
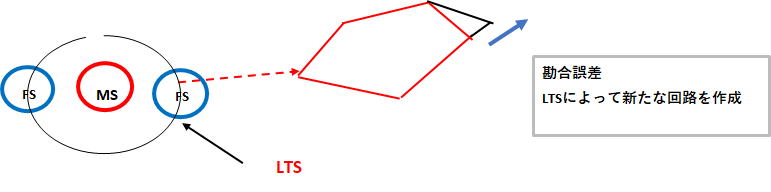
勘合システムにより照合されたお互いの描画は3次元であり高さの調整はドパミンとアセチルコリンで行うことは前述しました。次は形の調整ですが、これは第二の介在ニューロンであるLTSによるものと予想します。LTSは低閾値スパイク(Low-Threshold Spike)の略です。LTSによって二つの描画を合わせる。つまり該当箇所のMS(中型有棘神経細胞)を選択し、新たな神経細胞の組み合わせ(セルアセンブリ)を作る作業を行うのだと思います。
抑制の介在ニューロン(FS)に対して、抑制の介在ニューロン(LTS)の作用で興奮性の神経伝達が生じ、回路形成が可能になります。
(膜電位の閾値超え)
さて以上の作用により、描画12の高さと形の調整が終了して、その内容で行動命令が発せられるのですが、最後にもう一段の調整が入ります。それは、調整済の描画を構成するすべての神経細胞が発火する必要があるということです。つまり、膜電位が閾値を超える必要があります。

図1にそって説明します。まず用語の説明から。
① up & down とは特定部位の脳神経細胞の膜電位が定期的に上下する現象です。これは脳神経の自発的な発火により同期する神経集団が生じ、その影響が外部の脳神経に伝わる現象です。up state とdown state で10mvほどの膜電位の差が生じます。
② 大脳皮質から線条体へ行動命令が送られる際に、その情報はセルアセンブリであり同期しています。これが中型有棘神経細胞(MS)の周辺の 介在ニューロン(FS)に投射され、FSは同期して入ってきた情報をMSに伝達するとされています。
このように大脳皮質からの情報はFSにて選択され不必要(同期してない)な情報はオミットされます。ところが最近の研究により、
③ 大脳皮質から直接MSへ投射することが有ると確認されています。
さて、勘合システムで描画された図形(膜電位に差がある)は一括して脱分極(発火)する必要があります。というのは、描画形にこそ情報が統合されていて、個別の発火では意味をなさないからです。それでは、この一括発火と前述の①~③はどう関わっているのでしょうか。
これを個人的にまとめると次のようになります。
1 down
state このタイミングで大脳皮質からの行動命令をFSがMSへ伝達。同時に視床からの情報
と照らし合わせる勘合を行う。
2 大脳皮質から直接MSへ投射して膜電位を揃える。
3 up state で描画は持ち上がり、全ての神経細胞(MS)は脱分極(発火)する
4 大脳皮質から同期して入ってきた行動命令は、モールス信号的な情報伝達手段に変換され、淡蒼
球内節(外節)に流れていく
※ 青崎先生講義録 「線条体ニューロンの局所神経回路とその働き」
★ 閾値は固定して考えていて、実際は閾値は細胞内外の陽イオンの濃度で変更します。細胞内の陽
イオンの濃度はイオンポンプとイオンチャネルの働きによっていて、その動力源はアストロサイ
トからのATP供給によっています。
(勘合システムは存在するか)
さて以上で、私が予想するCSTC回路翻訳システムの説明を終えます。この予想が正しいかどうかは、ひとえに勘合システムが存在するかどうかであり、「線条体は大脳皮質からの行動命令と同時に視床からも同質の行動命令を受け取っているか否か」に拠っています。
これについては、私が思いついたわけではなくて、いくつかそれらしい論文があり、これは勘合だなと感じていた時に、視床線条体投射とストリオソーム・マトリックス構造 北海道大学解剖学分野組織細胞学教室 の論文を見つたという次第です。下記はその一節です。
「その結果、束傍核はマトリックスに優位に、正中核群からはストリオソーム優位に、束傍核以外の髄板内核群からはストリオソームとマトリックスに同程度の投射があることがわかりました。さらに、ストリオソームやマトリックスに特異的に投射する視床亜核の大脳皮質への投射先は、その視床亜核が投射している線条体のコンパートメントに優位に投射している皮質領域であることがわかりました。つまり、線条体のストリオソーム・マトリックス構造は、視床と大脳皮質から、時間差で同質の情報を受け取っている可能性があることが示唆されました。」
これを見た時、まさにその瞬間に、勘合システムの存在を確信しました。それなら「線条体は視床と大脳皮質から同質の情報を受け取っている」として、その後の処理はどうするのか。「必ず両者に誤差がありそれを埋める処理をするはずだ」となり、それが「線条体の皮質からの行動命令翻訳システムの中核になっているはずだ」と考えたのです。